(1). the capacity to meet novel situations, or to learn to do so, by new adaptive responses and
(2). the ability to perform tests or tasks, involving the grasping of relationships the degree of intelligence being proportional to the complexity, or the abstractness, or both of the relationships".
While Arthur S. Reber (1985) defines insight as:
"The process by which problems are solved. In this sense, insight characterizes a sudden re-organization or restructuring of the pattern or significance of events allowing one to grasp relationships relevant to the solution".
In that the clarification of experiences-or-adaptation of techniques to environmental circumstances (collectively abstractions, representations, organizations, etc) involves insight (after Wolfgang Köhler (1925), sudden re-organizing [re-representing] realizing, etc.; further higher order abstracting), the formation of a gestalt. In other words insight('feeling')-logic(Greek logike, art of 'reason': inferring, further the rules it operates by; comprises of 'deduction [Latin dēdūcere, from Greek apagogic, to lead: to 'deduce'; to infer necessary consequences from given premises, hence a priori from Latin for prior to observation(s): 'deduction' an 'identification' from 'universal' abstraction(s) to observation(s)] along with induction [Latin inductiōnem, from Greek epagoge, to lead: to adduce; to infer premises from given observations, hence a posteriori from Latin for after observation(s): induction from observation(s) to abstraction(s)])-about-event(s) (event-insight-logic; evaluating), in that insights become equivalent to jumps in perceiving (whether taken as senses or-both understanding) directing 'reasoning' in a process in transacting with event(s).
Now an insight otherwise an accumulation of insights would expectedly lead to noticeable progressive changes in organization: socially, technologically, scientifically, etc. Not only taught then passed on from parents to their offspring, but may involve subsequent improvements upon in successive generations. This insight along with the consequences, Korzybski (1921) refers to as Time-Binding: capacity to improve on the accumulated abstractions of others, then transmitting it for future generations, for example humans. Functionally distinguishing humans form plants (Chemical-binders: capacity to convert energy [for example, photosynthesis] into growth, etc) along with animals (Space-binders: move to find food, etc).
As Korzybski (1921) puts it in "Manhood Of Humanity",
"...This time-binding faculty is a functional name and definition for what we broadly mean by human 'intelligence'; which makes it obvious that time (in any understanding of the term) is somehow very closely related to intelligence".
Finally Korzybski provides a mathematical formulation of time-binding:
PRT Equation 1.
after Alfred Korzybski (1921). Such that in total gain in T successive generation:
R
— (PRT – P) Equation 2.
R - 1
after Alfred Korzybski (1921), in "Manhood Of Humanity".
where P, Progress made.
R, Ratio of improvement.
T, Generation.
Of course time-binding should not be special to humans alone, but can characterize any other forms of sentience. Korzybski (1933) provides differences that we might expect between animals with humans, hence between those sentient with those not:
(i). use of language;
(ii). a more developed cortex;
(iii). continued development of cortex after birth;
(iv). scientific activities;
(v). use of aids for recording or-both copying
symbols, for example books, computers, etc.;
(vi). etc (technology, etc).
Yet though the signs of progress, such as for example libraries, industries, etc., may not be present, particularly in young flourishing civilizations. Further as Korzybski (1933) points out:
"We should notice also that in the case of primitive tribes which apparently have not progressed at all for many thousands of years, we always find, among other reasons, some special doctrines or creeds, which proclaim very efficiently often by killing off individuals (who always are responsible for progress in general), that any progress or departure from 'time honoured' habits or prejudices 'is a mortal sin' or what not".
Nor in the case of Cetaceans living in the oceans may we expect (if ever), any kinds of constructions such as dwellings, factories, etc. What we can expect however, involves an increasing complexity of language, modification of tools-techniques, along with some form of social organization (relationships, etc). These subsequently become passed on as knowledge (experience) to successive generations, which may become further improved upon.
Therefore of the possible components of sentience, we have so far defined abstracting as apart of our definition of consciousness of abstracting. The expected using of tools, otherwise the degree of technological advancement, definable as for example by Swannell (1989) as "the study or use of the mechanical arts and applied sciences", otherwise more clearly as tool-use. Though as Carl Sagan (1992) argues, a distinction between tool using along with tool making appears appropriate. However, humans are not the only species who manufactures both uses tools. Because, as Sagan further observes so do Chimpanzees, for example in "fishing" for termites. As Jennifer Lindsey (1990) describes, from Jane Goodall's research:
"Chimpanzees use many objects as tools, and also modify them if necessary, such as grass stems, or twigs stripped of leaves, to extract termites from their nests.
They use leaves to wipe dirt off themselves or, crumpled as sponges, to sop up water they cannot reach with their lips.
They use rocks and branches as weapons during aggressions". Further their ability to use symbols appears a special case, like humans before they began their journey to sentience. Further social-organizing which appears implied by the process of time-binding as co-operating, can become defined as showing complex kinds of relationships between individuals in a group(s); as observed-inferred by Kurt Lewin's (1948) group dynamic social-field theory.
To these five introduced, two more possible candidates appear mentioned near the beginning. We have self-sacrifice (self-motivating), defined by Swannell (1989) as "the sacrifice of one's interests and desires to those of others". Where any evidence should constitute the willingness of an unrelated member of a species to show altruism, hence sacrifice themselves for another. Finally creativity, definable by Swannell (1989) as "inventive, imaginative", etc. Though this process involves insights concerning representations, it becomes continued for the beauty, one's own satisfaction, etc.; where evidence could constitute, art, fiction, play, etc., found specifically amongst the adults of the species.
Therefore sentient life entails a gestalt (packet; over-lapping because they are not 'elements') of:
(1). Technological: degree of tool-use-
manufacturing.
(2). Creative: innovative (Latin innovus, make new:
to alter, change, modify, introducing something
new, etc), imaginative, etc.
(3). Social-organizing (co-operating): showing
complex relationships.
(4). Abstracting: representing, gestalts, etc., of
noumena.
(5). Consciousness of abstracting: gestalt based on
the process of representing-as-a-whole from
noumena otherwise self.
(6). Time-binding: insights, the high order
abstractions made upon abstractions passed on
from previous generations.
(7). Self-sacrificing: altruism.
Ofcourse having made the case for sentience involving the above components. We must however mention that not 'all' humans have an intellectual-physical development equivalence. Apart from children along with the mentally-physically impaired, there remain those who cannot be relied upon to make (if ever) meaningful insights from accurate abstractions (observation(s), inference(s), etc), due to their 'Aristotelian-conditionality': education in 'Aristotelian methodology-or-logic' ('A', 'Aristotelian thinking'; methodology [principles concerning procedure for investigation, verification, etc., along with those of evaluating; hence a system for organizing knowledge] organizing knowledge as an 'intensional' [false-to-facts 'universalizing'] language 'habit(s)' based on the laws of 'identity' ['everything is the same'], excluded third ['or not'], with contradiction' ['but not both'], systematized by Aristotle (350 B.C.), 'dogmatised' by his followers), hence equivalent to 'identification'. For the description of time-binders, really only involves a few in any society over space-time. Specifically many scientists, engineers, further throughout recorded history only a handful of 'philosophers'. Such that through these components may not appear to apply to many human adults (let alone children), should not be a paradox, since sentience remains as a result of an evolutionary (adaptation) process.
For example a similar problem does occur with the characteristics of life, which M.B.V. Roberts (1971) gives as:
(1). Movement.
(2). 'Responsiveness' (sensitivity otherwise irritability).
(3). Growth.
(4). Feeding.
(5). Reproduction.
(6). Release of energy (respiration).
(7). Excretion.
For if a human adult does not get to reproduce themselves during their life, does this mean these specific human individuals are not life-forms. Instead the characteristics of life represent average expectations (that as such if alive, then such organisms have these characteristics as capacity) of life-functioning-as-a-whole.
Therefore while many human adults along with children, may not be sentient according to the criteria, such that these human adults may become more accurately described as infantile. The characteristics of sentience functionally describe a species-as-a-whole, not any single organism.
Whereas those 'species' that fulfil some of the conditions, may co-incidentally do so anyway, for example growth appears a characteristic of life, yet crystals of non-living materials grow. Indeed animals will usually only form 'identifications' as abstractions. Alfred Korzybski (1947) discussed experiments further exploring the mechanism of 'identification', as a form of abstraction (representation), in dogs. He found that though dogs can form a number of abstraction levels, yet because animals do not have Science (Latin scientia from scīre, to know: knowledge attained by experiment-observation-induction), etc., they do not know - cannot know they abstract, hence have not benefited beyond basic survival terms. Upon Pavlov's (1906) experiments, Korzybski (1947) reported that dogs appeared capable of forming 2-4 levels of 'associative' abstractions, where Pavlov's bell for food represents the 1st level. A 2nd order abstraction appears possible when the dog comes to 'associate' a whistle with a bell, after 'associating' the bell for food. However this appears limited to only two levels of abstraction for food, since when if one attempted to 'associate' a metronome with the whistle, the dog did not come to salivate. But in terms of warning against 'pain', for example acid in the mouth, the dog can abstract to a 3rd level by 'associating' a metronome with the whistle. Further W. Horsley Gantt found that in terms of 'sex' (the biological imperative), dogs can form a 4th level of abstraction. Thus in survival terms, 'sexual urges' appears stronger than 'pain', while the avoidance of 'pain' more important than the attainment of nutrients. However, as Korzybski (1947) argues, since animals cannot know they abstract, then the number of ('associated') abstraction levels must finish at some point. However, this kind of explanation as established in Pavlov's experiments, where something 'is always' expected to follow, has an equivalence to an (mis)evaluation in humans which nevertheless involves an 'identification' as discussed.
Now of the various candidates mentioned, Bottle-nose dolphins have perhaps attracted the most research so far. The brain of the Bottle-nose dolphin appears very large, which Pilleri (1968) gives as weighing 2.3Kg (5lbs). Having highly convoluted (folded) cerebral hemispheres, of which the neo-cortex forms 98 percent though thinner than with humans. Though this large brain appears only as indirect evidence for sentience, nevertheless the differences in the structure to that of our own brain would suggest a different from of sentience, created through evolution in a different environment. Now we must prove that this large brain despite differences, has recognizable functions in terms of our criteria for sentience. Therefore let us test whether Bottle-nose dolphins fulfil our established criteria for sentience.
Evidence for abstracting (4).
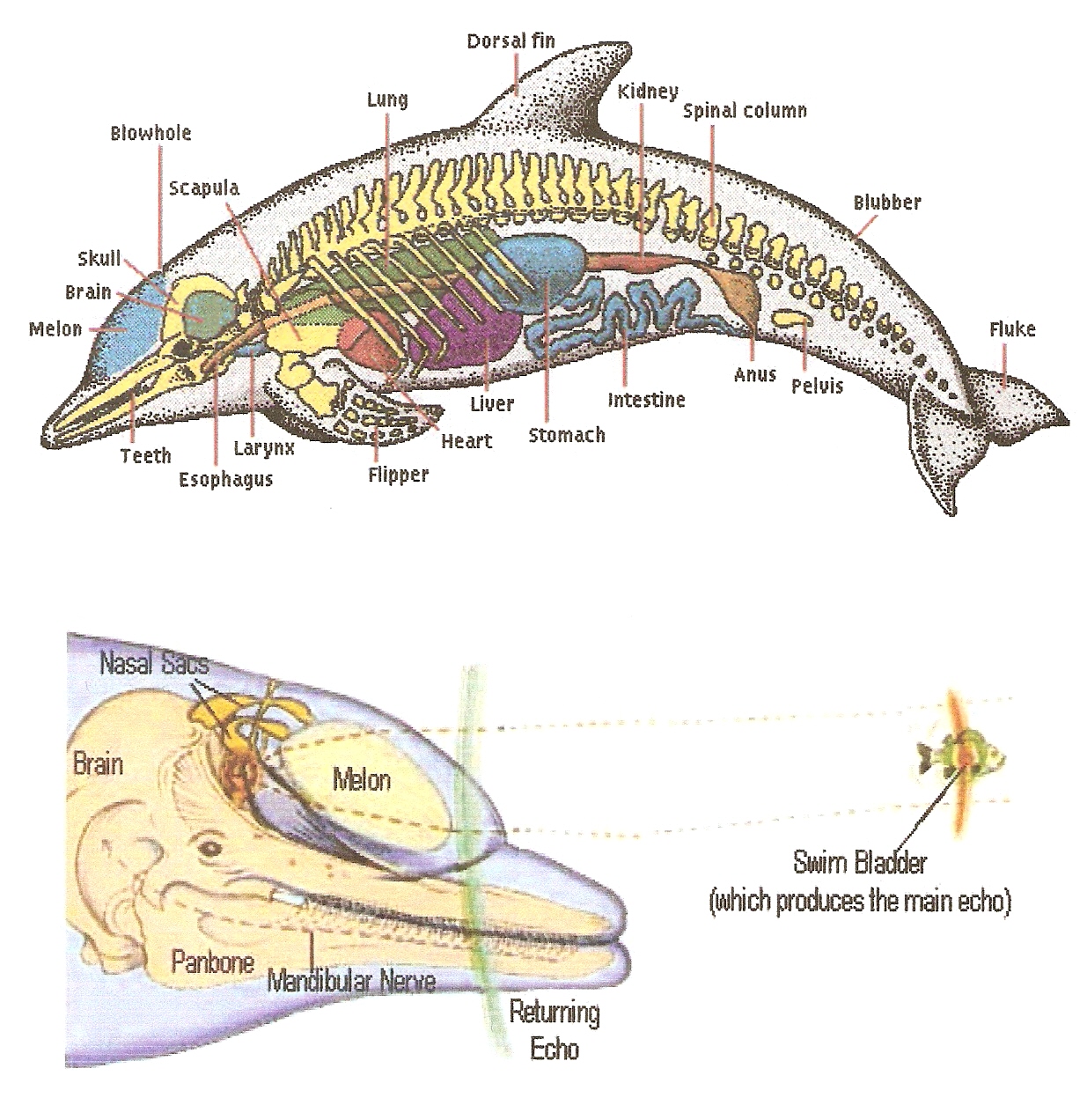
Bottle-nose dolphins remain capable of forming representations involving the perceptions of 'sight', 'touch', 'sound', along with 'taste-smell', where there is not a clear distinction between these last two in the medium of water, further they may 'sense' the earth's magnetic field. 'Taste' buds at the base of the tongue along with nerves going to the brain indicate that dolphins experience some sort of 'taste perception'. Experiments on Bottle-nose dolphins have shown that they can distinguish chemicals, such as different concentrations of citric acid as sour-bitterness, but appear poor at detecting salty-sweet 'tastes'. Whereas dolphins have little else no 'sense of smell', as 'olfactory' nerves with lobes in the brain appear absent; where the nostrils for example have developed into a blow-hole. As a result whether pheromones appear important to indicate sexual-social dynamics, though secreted (here released in water), remains unknown. 'Touch' appears surprisingly well developed, where a dolphin's skin appears tremendously 'sensitive' around the head, dorsal fin, flukes along with flippers, used to detect water currents, etc. Further dolphins frequently 'touch-caress' each other with their pectoral flippers along with their genital organs. A dolphin has excellent eye-sight above-both-below the water surface because the lens along with cornea can correct for the transition between water with air. The light-sensitive retina at the back of each eye contains rods-both-cones, so that a dolphin can see well in bright-both-dim-light, further in color. Further a tapetum lucidum, the mirror-like light-reflecting layer allows more light through the retina, providing enhanced 'vision' at low light levels. Two central faveae (light-sensitive areas on each retina, of which humans have only one) provide dolphins with nearly 3600 'vision', with a narrow forward-facing field of binocular 'vision' to the front; though a blind spot directly behind. Nevertheless 'vision' becomes limiting at depth (where light increasingly diminishes), obscured conditions, etc. Therefore 'hearing' has become the main sense of dolphins. The most basic aspect of their hearing involves their ability to make use of noises currently in the environment. The hiss of sand swept along the sea bed may indicate where currents appear strongest, whilst the roaring of swells otherwise frizzing of rain up above somewhat determines how the dolphin might best surface. Their prey make noises too, many bottom-living fish make grunting calls, etc. However even more complex, a dolphin has the ability to make-receive noises of their own. Dolphins click, bark, yelp, bray, pop, whistle, mew, squeal, buzz, etc., as if torturing a variety of farmyard animals. Furthermore, the dolphin cacophony extends way above our own hearing range into ultra-sonic frequencies.
Despite considerable study, the functions of most of these calls are still not known. By far the most famous involve echo-location, which allow dolphins to detect objects in the water in front of them (though there remains a blind spot behind). Lyall Watson (1981) reports upon an observation made by Arthur McBride in 1967 published in a paper by R. McNally (1977):
"In 1967 Arthur McBride curator of the Marine Studios in Florida, discovered that Bottlenose Dolphins (Tursiops truncatus) were able to evade his nets even where visibility was virtually nil (McNally 1977). He theorized that they were using a system of echolocation similar to that discovered in bats. In 1956 William Schevill and Barabara Lawrence published the first proof of this talent in Bottlenose Dolphins which produced creaking sounds while locating fish held silently under water".
They do this by making extremely loud clicks, then interpreting the echoes that return. A review of the data, suggests that the dolphin tunes their clicks to obtain the best possible information. It takes about six clicks to optimize the signal from the interrogated object. First the dolphin sends out a multi-functional click, then depending on what it has found, for example fish, crab, boat, diver, shark, etc. It modifies the next click, the clicks 'settling' after about half a dozen. The dolphin will not send out a fresh click until it has determined the significance of the previous one, using this information to modify the next click. The process however remains rapid, such that a searching dolphin will sound much like a slowly revolving bicycle wheel. These clicks remain very high frequency sounds, beyond the reach of human hearing. While the hearing range of dolphins appears as 1-150KHz (compared to a human range of 0.02-17KHz), where peak sensitivity has an equivalence to 40-100KHz. Dolphins can hear sounds with frequencies below 1KHz, if they become loud enough, further there remains speculation that tiny external ear openings just behind the eyes can receive sounds below 20KHz. We now know that the dolphin's echo-location clicks, vary depending on the species that makes them; Bottle-nose dolphins produce clicks that span a wide range of frequencies. Though only a few Odontocete species have become experimentally proven to have the ability to echo-locate. However the anatomical evidence, the presence of the melon, nasal sacs, specialized skull structures, etc., suggest that dolphins-as-a-whole have this ability. Refer to Figure 26. The dolphin generates the sound in the form of clicks within the nasal sacs, situated behind the melon. The frequency of this click differs as to function, further differs between species. The melon functions as a lens which focuses the sound into a narrow beam projected away out in front of the dolphin. When the sound strikes an object, some of the energy of the sound-wave gets reflected back. It then appears that this echo gets received by the panbone, whilst the fatty tissue behind it, transmits the sound to the middle ear, thence to the brain. Further the teeth of the dolphin along with the mandibular nerve that runs through the jawbone, may become involved. Using only their acoustic-sense a Bottle-nose dolphin can discriminate between objects which differ by ten percent perhaps even less, in volume otherwise surface area. As Watson (1981) elaborates:
"The denser the medium, the faster sound waves travel in it. The speed of sound in water is roughly four times as high as it is in air. The distance a sound travels in either medium depends on its wavelength or frequency. High-pitched sounds travel only a fraction as far as low-pitched sounds of the same power. So, for long-range communication most animals use low-frequency, long wavelength sounds like the 20-Hz trans-oceanic moan of the Fin Whale. But a 20-Hz signal has a wavelength of almost 75 m (250 ft), which means that it will pass unimpeded over most obstacles, bouncing back only off something like an oil tanker or a seamount which has a diameter greater than 75 m (250 ft). The sonar systems of toothed whales concentrate on the short-range higher frequencies. Dolphins in tests locate fish less than 20 mm (.75 inch) long, so they must be using sound of a frequency near 100 kHz. Some cetaceans produce sounds with, and are presumably equipped for detecting the echoes from, frequencies higher than 200 kHz, with which they can distinguish objects as small as vertical wires 0.35 mm (less than .015 inch) in diameter (Penner aswell as Murchison 1970).
Most toothed-whale sonar systems operate with a steady stream of intense click sounds, generally between 30-60 kHz. They can vary the rate of clicking (up to 1,000 per second) for building more accurate echo pictures and can change the frequency of their transmissions, moving the energy peak up and down. Some can even produce two different frequencies simultaneously, and the most efficient of these can flatten, shorten or heighten the sound beam in order to focus it on a target. A few can manipulate sonar probes precisely enough to send a secondary exploratory beam off at right angles to the main one (Evans 1973).
Echolocation in toothed whales fall into four categories:
(i). Low-frequency, narrow band, with peak energy
below 4 kHz (e.g. The Common Porpoise).
(ii). Low-frequency, broad band, with peak energy
between 16 and 20 kHz (e.g. the Great Killer
Whale).
(iii). High-frequency, broad band, with peak energy
between 30 and 60 kHz (e.g. the Bottlenose
Dolphin).
(iv). Ultrasonic, broad band with peak energy above
80 kHz (e.g. the Indus River Dolphin).
These species also have lower-frequencies sounds for communication, and some of them may operate in more than one of these modes, depending on what they are doing. All
Figure 25-6.
Anatomy of Tursiops Truncatus.
of them achieve greater acuity in any mode by 'scanning', by sweeping their sonar beams over the target with movements of their heads or bodies as they listen. This also changes the angle at which the returning echoes strike the head, and therefore the degree to which each frequency penetrates and is heard, providing very precise information about the nature of the object".
Michael Bright (2001) reports that the echo-location system appears so effective, that a Bottle-nose dolphin in captivity has distinguished between a tangerine with a small metal ball, at a distance of 113 metres; sounds so powerful that in water can travel close to the speed of sound. For example Watson (1981) reports on an experiment made by William Evans (1966):
"After training a blindfold female Bottlenose Dolphin (Tursiops truncatus) to recognize a copper plate by its acoustic properties, William Evans of the Naval Undersea Center in San Diego presented her with glass, plastic and aluminium imitations of the same size and shape. She immediately dismissed these as counterfeits. He then tested her with an aluminium plate whose thickness had been calculated to have exactly the same sonic reflectivity as copper, but she still picked out the copper correctly. To do this she had not only to perceive the basic features of the echoes but also their precise frequency composition. What makes this performance even more impressive is that, to do it at all, she had to remember the frequency mix in the copper echo, analyse the sound from the aluminium copy, and then compare the two for subtle proportional differences. She completed this sophisticated exercise in a matter of seconds (Evans & Powell 1966)".
Indeed a dolphin can echo-locate in noisy environments, on near or-both distant targets while further whistling, etc., feats which have engineer-scientists in owe.
Bryde with Harrison (1988) noted that when a captive Bottle-nose dolphin became taught to attack sharks. The Bottlenose dolphin would attack Sandbar sharks (Archarhinus Plumbeus), Lemon sharks (Negapnon Brevirastrics) along with Nurse sharks (Ginglymastama Cirratum), none of whom have been known to attack dolphins. However, the Bottle-nose dolphin refused to attack a Bull shark (Carcharhinus Leucas) a known predator of dolphins. Bryde with Harrison suggests that this involves Bottle-nose dolphins in 'classifying' (Greek katēgoria, to 'categorize': process to order-or-group by 'two-valued, el, allness by same properties' from Greek proprium: that owned by; typical 'element', etc.; equivalent to 'identification(s)') species of sharks according to the threat that they may present.
Captive studies have further tended to concentrate on the dolphins capacity for language. Douglas Richards, James Wolz with Louis Herman (1984), studied vocal mimicry by training Bottle-nose dolphins to mimic computer generated model sounds by modifying their whistles. They found that modelled sounds not heard previously, could become mimicked immediately with high reliability in repeated tests. In further trials involving 'visual' memory, dolphins came to use whistles as vocal labels for objects. But appear to under perform until they could relate to a particular object (for example ball, hoop, etc) to a presented sound. Further suggesting the importance of 'auditory' representations have, in making 'sense' of their world.
Experiments by John Cunningham Lilly (1978), Wilhelm Dudoc Van Heel, etc., have succeeded in getting Cetaceans to acknowledge human vocalizations, for example whistle language, words, etc. Indeed one of Lilly's (1965) assertions involved dolphins having the capacity to communicate with humans, mimicking human speech. Lilly along with colleagues placed captive Bottle-nose dolphins in an experimental chamber with their blow-holes exposed, to whom a long list of human speech sounds, both as random phonetic units along with in the form of simple English words-phrases. Fish rewards got offered to the dolphins in order to encourage them to produce more air sounds. The dolphin compared to human sounds showed many similarities, producing sounds quite unlike their own natural ones, in the frequency, number, along with gaps between bursts of sound involved. They came to mimic their human experimenters. However Lilly's opponents have strongly criticized the dolphin 'responses', questioning their alleged 'voice and accent' along with their comprehension. Though originally discounted, Bright (2001) has since reported that dolphins have indeed learned to count in English, further appearing to comprehend the meaning of the simple phrases they can speak, such as 'stop it', 'more', 'bye-bye', etc. As Bright (2001) recounts:
"In 1963 two scientists peered over the edge of a tank at the communication Research Institute on St Thomas in the US Virgin Islands. Elvar the dolphin, spotting at least one familiar face, swam over and presented his belly at the surface. The animal was inviting the scientists to tickle him. One scientist obliged and scratched him several times, but on each occasion the dolphin rested a little deeper in the water. At last, the dolphin was too deep to reach and the scientist gave up, removing his arm from the tank. Instantly, the dolphin swam to the surface, raised itself up until it stood on its tail and, towering over the two men, clearly enunciated the word 'more'. Had he understood the meaning of 'more' or had he simply mimicked a sound that triggered the desired response? Either way, it was a momentous occasion for astronomer Carl Sagan and pioneering dolphin researcher John Lilly".
Herman's (1980) group further taught artificial languages to two Bottle-nose dolphins. One of them a special form of sign language based on gestures, the other an acoustic based language. These studies intended to test for sentence comprehension, rather than language production. To this end, various objects (such as balls, pipes, frisbees, hoops, etc) placed in their pool became presented so that these related to a sound, otherwise a gesture. They then become taught a variety of verbs, for example 'touch', 'fetch', etc. Which then got introduced into simple two-three word command sentences, for example once the dolphin has learnt to fetch a ball, it may get asked to take it to the hoop by saying "ball-fetch-hoop". Herman's (1984) extended studies, have shown that dolphins can understand command sentences up to 5 words long, for example "touch the ball with your tail". Coming to understand qualifiers such as up, down, over, under, through, left, right, etc. Distinguishing orders of words, for example "ball-fetch-hoop" with "hoop-fetch-ball". Robin Williams (1997) in the documentary "Dolphins With Robin Williams", reported that Botttle-nose dolphins could comprehend around sixty words forming around 2000 sentences. As a result these Bottle-nose dolphins have demonstrated an understanding of semantics (Greek semainein, 'meaning'), syntax (Greek suntaxis, to arrange: part of grammar concerning the arrangement-or-construction of sentences), etc., the rules underlying language. Bryde with Harrison further report that Herman's group, found that these studies include that Bottle-nose dolphins can form higher abstractions such as the rules underlying the 'behavior' that they may carry out. While Herman with P.H. Forestell (1985) have noticed, that language-trained dolphins do know if an arbitrary object appears absent from their environment, since they report it.
Further David Attenborough (2005) in the documentary "Wildlife On One: Dolphins Deep Thinkers?", reported on extended experiments conducted at the Marine Mammal Laboratory in Hawaii, on dolphin 'reasoning', involving three Bottle-nose dolphins: Phoenix, Akare-cumy, along with He-a-po. The first two experiments demonstrated that dolphins can distinguish between objects whether via echo-location, vision, or-both.
Using a plastic cross, metal bowl, otherwise a ceramic flower-pot presented out of sight of the dolphin in a sample box (over-hung wooden framed box, partly immersed in the pool), the dolphin then after using echo-location, had to select the correct object held-out by other researchers at the far-side of the pool. A task which He-a-po performed flawlessly.
In the second a researcher would hold out two objects, for example two flower-pots, with which the dolphin had to decide whether the objects appeared similar otherwise different, by selecting one of two poles held out by other researchers. Remarkably even when Akare-cumy got presented with objects she had not seen before, she still discerned any similarities-differences by selecting the correct pole.
Dolphins can mimic 'actions', after the first occasion they see it. For example Akare-cumy can relate her own 'body' parts to the researchers, for example fins for hands, tail for feet, etc. Imitation appears useful in learning from others, for example calves imitate their mothers. For example, Bottle-nose dolphins in an Australian bay, chase fish to force stranding on the beach in shallow water. This however requires two difficult techniques: hydro-planing along with un-beaching themselves. Both requiring practice, experience, etc., particularly of the calves who begin by mimicking their mothers. However when Bottle-nose dolphins of the tidal-creeks of South Carolina US, co-ordinate their movements in beaching (on their right-side) to catch the forced stranding of fish, yet how does innovation, mimicry along with co-operation relate? In an experiment, when Phoenix along with Akare-cumy got given a tandem sign-instruction: "kiss-jump together", they then went off to perform in unison. Yet when given the tandem sign-instruction: "create", both perform as if in a synchronised routine, one appearing to take the lead (as in kiss-jumping earlier), while the other copies. However examples of inventiveness include bow-wave riding, surfing, etc.
However over the years the Hawaii researchers have developed a sign-language, in order to communicate with the dolphins. For example, Akare-cumy has learned a vocabulary of over 60 words including squirt, person, etc., even a sign for her friend Phoenix; to do so, she has ofcourse to understand what each sign refers to, along with how order contextually changes 'meaning'. In order to test this, researchers using blacked-out goggles (preventing cues via eye contact) would give sign-instructions to the dolphins. Such that, when Akare-cumy got the sign-instruction: "Phoenix-pipe-fetch" (take the pipe to Phoenix), Akare-cumy showed that she understood that Phoenix referred to her play-mate. While in order to test word order significance, such sign-instructions given include: "right-water-left-Frisbee-fetch" (take the Frisbee on the left, to the jet of water on the right), "left-water-right-Frisbee-fetch" (take the right Frisbee, to the left jet of water). Which again Akare-cumy demonstrated a flaw-less appreciation.
Indeed dolphins can understand the significance of referring to objects by pointing; further this referring to objects can become achieved by just gazing at it. However 'pointing' (gesturing at) though appears simple, it can take children up to a year to develop this skill; whilst Chimpanzees appear one of the few other animals that may appreciate what a point 'means'. Experiments demonstrating this appreciation with dolphins, involves the researcher pointing at one of the presented objects, then continuing to sign-instruct as usual. For example, after pointing at an object such as cross, bowl along with flower-pot, the dolphin would then have to find a match at the other end of the pool, where other researchers would hold out similar objects.
In order to ask the dolphins questions, researchers had the notion of introducing two paddles: one a white flat-paddle 'meaning' "yes", the other a black rubber cap on a short pole 'meaning' "no". Then a researcher would ask a dolphin some questions, with which involving the paddles the dolphins can answer yes-or-no. For example, after presenting a pipe-and-basket to Akare-cumy, the researcher asked: "is there a surf-board in the pool?", otherwise "is there a pipe in the pool?" Of which Akare-cumy answered correctly via the appropriate paddle.
However when given a trick sign-instruction: "hoop-basket-fetch" (take the hoop to the basket), while there was no hoop in the pool. Akare-cumy ingeniously collected the basket, then pressed the no-paddle, surprising the researchers.
Further David Attenborough (2005) took part in one experiment demonstrating that dolphins can understand the significance of television. Since a dolphin will still follow sign-instructions given by Attenborough while appearing on a television screen, presented at a viewing port in the side of the pool. The sign-instructions at first involved simple ones, for example "blow-bubbles", "roll-over", "butter-fly swim", etc., including imitating head-nodding up-and-down; progressing to "Frisbee-basket-fetch" (put the Frisbee in the basket).
Such that these studies, observations, etc., show that Bottle-nose dolphins not only appear able to abstract, but further capable of higher order abstracting; possible only if free from 'identifying'.
Though these studies remain exciting, it would appear more convincing if dolphins in their natural environment showed an equivalent capacity for abstracting. Indeed as far as gestures go, posture, head turning, 'touching', even bubbles exhaled (for example, in sound-making, surprise, etc), etc., these do play an important part in communication; though these do require clear environmental conditions for their perception.
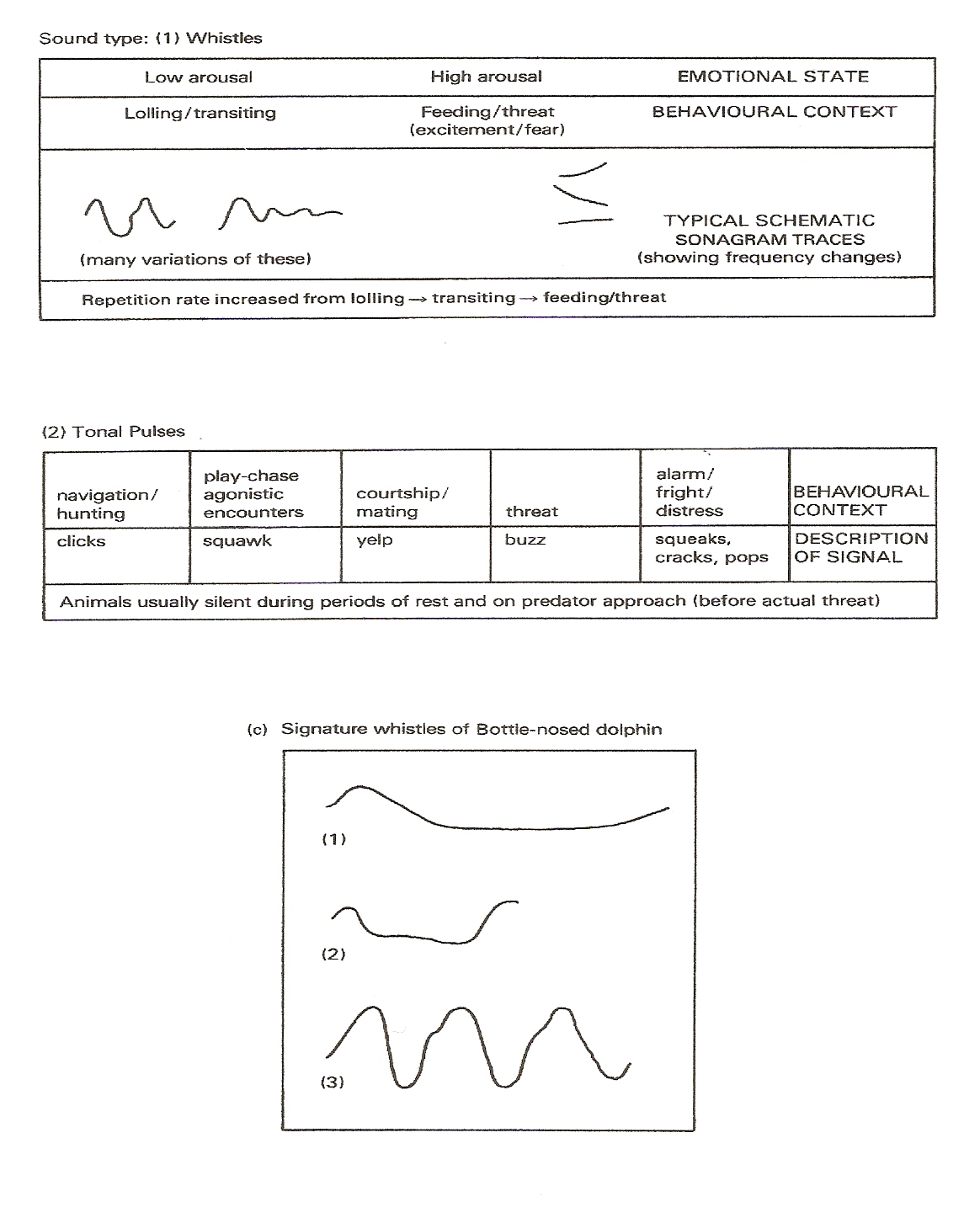
Now as mentioned the vocal repertoire of dolphins clearly varies, containing squawks, squeaks, whistles, burps, groans, clicks, buzzes, barks, rattles, chirps, chuckles, moans, etc. Yet they appear grouped roughly into pulsed sounds involving ultra-high-frequency clicks, burst pulses including click-trains, chirps, yelps, chuckles, raspberries, etc., (made up of rapid clicks) along with un-pulsed sounds involving frequency-modulated whistles, squeaks, etc. Many pulsed click-sounds have echolocation-navigation functions, but some further have a social function. Pulsed yelps can occur in courtship, pulsed squeaks during distress, buzzing click-trains during aggressive confrontation. However since high frequency clicks travel only short distances (echo-location remains highly efficient at ranges of less than 100 metres), then it appears that they become suitable for communication only within small groups in close contact with one another. Un-pulsed whistles instead carry better over long distances, so these might appear most useful in large groups.
Kenneth S. Norris (1992) reports on research performed at Sarasota bay Florida by Randall Wells' research group. There an experiment based on recorded whistles taken directly from a Bottle-nose dolphin's melon, showed that these become used as recognition for each other. Where these whistles got replayed to a second Bottle-nose dolphin known by the first, alternated with whistles of another dolphin though part of the community not so well known to the second. Evaluation of the video tape, data, etc., showed that the dolphin became more excited to the whistles from her friend.
Each dolphin develops their own distinctive signature whistle from an infant, which other dolphins come to recognize, rather like a name for each other. By listening for these particular whistles, a dolphin can keep track of the position of other members during travel-foraging, etc., when 'visual' contact is not possible. Further by imitation of another whistle, a dolphin can call a particular individual. Peter Tyack a colleague of Wells further speculated that:
"While some acoustic features of a signature whistle are stable others may vary. These changes may pass on additional information that all the other dolphins in the area can listen in on. This means that a society based heavily on learning can be maintained inside a dolphin school".
In other words dolphins may live in their own educated culture.
Jarvis Bastian (1967) carried out one of the early experiments to show that dolphins communicated information to one another. In 1965 he placed a pair of Bottle-nose dolphins in adjacent tanks isolated visually from one another, but in sound contact. The female then got taught to push an array of paddles for which it received a reward. The male got presented with a similar array of paddles but given no training. Yet the male eventually pushed the correct paddle. Bastian assumed that the female had passed on the necessary information to the male. However the information may have become passed inadvertently, otherwise the male may have worked it out independently.
More convincingly J.F. Fish with G.E. Lingle (1977) reported that captured a male Pacific Spotted dolphin (Stenella Attenuata) whistled virtually continuously, probably as a result of alarm-distress. When these sounds got recorded then played back to members of the dolphins school, they fled; yet when played to another school they showed curiosity. Presumably the dolphins own school had picked up a danger signal from the familiar call of the captured dolphin, whereas the strangers were not able to appreciate the significance of the call.
Other research (J.J. Dreher with W.E. Evans (1964): W.E. Evans with J.H. Prescorl (1962); M.C. with D.K. Caldwell (1979); along with Herman with W.N. Tavolga (1980)) has revealed that dolphins deliver their usual calls in a particular way to convey their 'emotional' condition. Refer to Figure 3. Indeed Bright (2001) reports that female dolphins in the wild-both-in-captivity, for example have become observed trying frantically to revive a dead calf. Evaluation of the calls from captive dolphins swimming normally, compared with those from dolphins removed from the tank for veterinary examination, has revealed that dolphins under stress emit calls that have a significant pitch or-both duration change. But apart from indicating that dolphins can communicate 'emotions', these studies further suggest that dolphins rely so much on their 'acoustic sense', that they may appear able to project their 'thoughts' in the form of 'acoustic' rather than


Figure 27-8.
Randall Wells' research group at Sarasota bay Florida. There an experiment based on recorded whistles taken directly from a Bottle-nose dolphin's melon, showed that these become used as recognition for each
other.
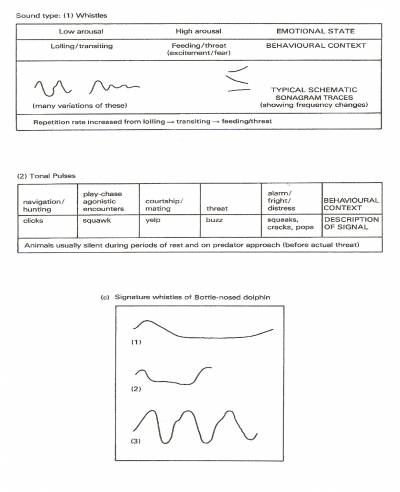
Figure 29.
Language in Delphinids: the contexts in which different calls get made by Bottle-nose Dolphins based on J.J. Dreher with W.E. Evans (1964); W.E. Evans with J.H. Prescott (1962); M.C. With D.K. Caldwell (1979); further data from Louis Herman with W.N. Tavolga (1980).
From Peter Evans (1987) "The Natural History Of Whales And Dolphins".
'visual' imagery, into the 'minds' of other dolphins in their groups. Further refer to evidence on tool-use (1) for understanding of causality, a higher order abstraction.
Evidence for tool-use (1).
Bryde with Harrison (1988), Norris (1992), documentary "Dolphins With Robin Williams" (1997), along with Bright (2001): each report on Bottle-nose dolphins on the Georgia coasts, South Carolina coasts, etc., of the United States, catching Mullet by using their tails to produce bow-waves, hence beaching the fish by mini-tidal waves. Then the dolphins launch themselves onto the beach, on their right side, grabbing as many fish as they can, while anchoring themselves with their pectoral flipper, before sliding back into the water. Refer to Figure 33. For example, as reported by Watson (1981), form Hoese (1971):
"Bottlenose Dolphins have been seen feeding in pairs in a tidal creek near Boboy Sound in Georgia (Hoese 1971). They come into the salt-water marsh there at low tide and, working as co-ordinated couples, rush at shoals of small fish in the shallows and sweep them with the force of their combined bow wave up on to a muddy bank. The dolphins then shoot out on to the mud slope themselves, sometimes coming completely clear of the water, snap up the stranded fish and slide back in".
Bright (2001) along with David Attenborough (2005) in "Wildlife On One: Dolphin Deep Thinkers?", further reports that Bottle-nose dolphins off the South Florida coast have developed a unique strategy in hunting Mullet. In shallow, muddy water, upon detecting a shoal of Mullet, a dolphin would break off circling the shoal in a ring ("curtain") of mud using the tail. Then the dolphins would line up on their sides, awaiting the panic-stricken fish to jump out of the water into their mouths. While further up the Florida coast, where though shallow, instead these shallows get covered in sea-grass, other Bottle-nose dolphins demonstrate versatility in 'behavior' by developing a different strategy. One dolphin using a thrashing tail, sends shock-waves through the water while circling the Mullet, forcing them to shoal together. Though again the remaining dolphins wait on their sides for the jumping fish.
These examples not only suggest an understanding of causality (as such evidence for abstracting), but that the tail to create a bow-wave, the use of beach, the creation of a mud-wall coupled with shallow water, appear as equivalent to tools - versatile-modify-inventiveness.
While Bright (2001) reports that female dolphins of Shark bay Western Australia, place sponges on the tips of their snouts. The sponges function to protect by covering the delicate snout when foraging among jagged rocks, corals, etc., against spines-both-stings of animals such as Stone-fish, Sting-rays, etc., hidden, when ploughing through the sand.
William Schevil with Barbara Lawrence (1956) published the first proof of Bottle-nose dolphins using echo-location to hunt, producing creaking sounds while locating fish in muddy water. Further Herzing (1996) while filming Atlantic Spotted dolphins (Stenella Frontalis) at North Bahamas off Florida, using echo-location while seeking razor fish, flauders, etc., noticed that the fish would go limp during their escape from beneath the sand. This led Herzing to the notion that dolphins not only use echo-location to find prey but hence to stun them too. Where such observations had previously led Norris with Bertel Mohl (1983), to propose their big-bang theory based upon further systematic investigation. They found that the sounds are not the very high frequency clicks usually used in echo-locating, but low-frequency 'bangs'. Such that dolphins can use these high-energy bursts to kill, if not then at least will result in debilitating prey at a distance by impairing their equilibrium (Latin oequilībrium, to balance) or-both sensory system, hence making them easier to catch. As a result any fish that have gotten zapped, swim in circles otherwise remain motionless. Norris (1989) along with colleagues further presented evidence involving experiments where they exposed Anchovies, to pneumatically-generated 'loud impulse sounds' similar to those recorded in the wild by dolphins. They discovered that these sounds killed otherwise injured the Anchovies.
Further Watson (1981) importantly relates:
"Our dependence on tools, which now extends to books and computers, means that a large part of the mental

Figure 30-2.
World-wide Dolphins employ many different strategies to catch fish, etc. A Bottle-nose Dolphin may search for food by echolocating near the sea bottom. As it emits echolocating sounds, it will move their rostrum (otherwise beak), back-and-forth, it may bury itself up to their eyeballs, practising "Crater Feeding", like those off the Bahamas, around Mikura Island, Japan. Dolphins may furthermore use their sounds to stun their prey.
From Tim Cahill (2000) "Dolphins".


Figure 33-4.
Bottle-nose Dolphins co-operate to feed on bait-ball; while it feeds the others patrol the sides to keep the prey tightly herded. From Kenneth S. Norris (1992) "Dolphins In Crisis", National Geographic.
Tim Cahill (2000) "Dolphins".
capacity of a human culture exists outside the individual, whereas in non-manipulative dolphins it is still embodied in brain and behavior. As far as our ability to act as social beings is concerned, we may be inferior to many cetaceans".
|